
研究
私たちは、神経細胞とグリア細胞、血管細胞等で構成される脳内の環境がどのように維持されているか、更にその破綻がどのようにして疾患の発症に結びつくのか、というテーマで以下の研究を行っています。
- 小胞体ストレス応答による脳内環境維持とその破綻
- アストロサイトによる脳内環境の改善
- 自閉症関連遺伝子による脳内代謝制御とグリアネットワークの形成
- 末梢細胞による脳内炎症の誘導とその制御
- ヒト脳の組織学的検討
以下、それぞれについて説明します。
1.小胞体ストレス応答による脳内環境維持とその破綻
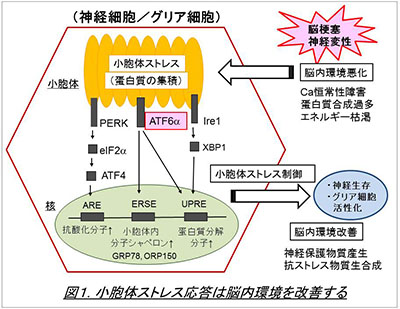
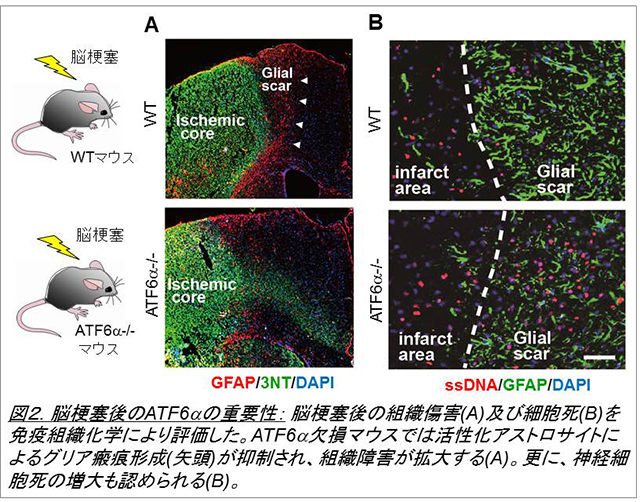
神経細胞は環境変化にきわめて弱く、例えば脳梗塞の際に、短時間で細胞死に至ると考えられています。一方、グリア細胞の一つであるアストロサイトはこの環境変化に対して強く、虚血環境でもかなり生き延びることができます。私たちはこのアストロサイトにおけるストレス応答に注目し、そのメカニズムを解析しました。そして、細胞内小器官の一つである小胞体におけるストレス応答が、脳環境の維持に重要であることを見出しました(図1, Tamatani et al., Nat Med. 2001, Hori et al, J Cell Biol. 2002)。小胞体ストレスは脳虚血(脳梗塞)以外にも、神経変性や外傷などの病態で発生しますが、これまでその詳細については不明でした。私たちは小胞体ストレス応答の主要3経路のうち、特にATF6α経路についてノックアウトマウスや培養細胞系を用いた解析を行い、同経路が神経細胞の生存(Hashida et al., 2012, Kezuka et al., 2016)を促進するほか、アストロサイトやミクログリア等のグリア細胞の活性化(図2, Yoshikawa et al., Ta et al., 2016)にも必要であることを明らかにしました。ATF6α経路は分子シャペロンの発現などを介して小胞体環境の改善に役立つと考えられており、今後は、本経路を利用することで、脳梗塞や神経変性疾患で起こる神経細胞死を抑制できないか、と考えています。
2.アストロサイトによる脳内環境の改善
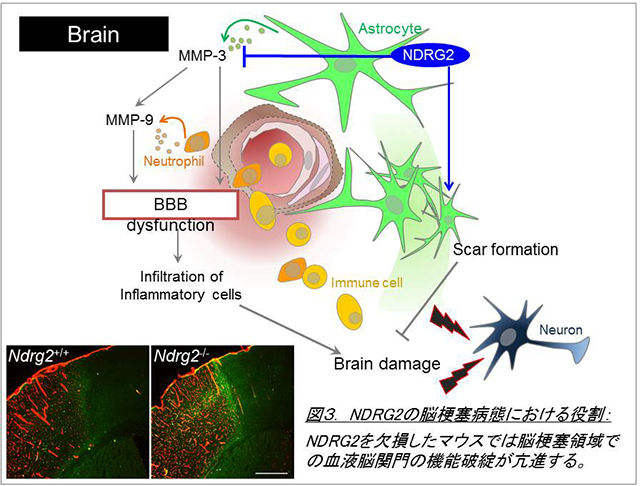
最近の研究から、アストロサイトは単に神経細胞を補助するだけの細胞ではなく、積極的に神経伝達や血流を制御し、障害時には脳環境の悪化に応答する極めて重要な細胞であることがわかってきています。私たちは中枢神経系でアストロサイト特異的に発現する新規分化関連遺伝子NDRG2 (N-myc downstream regulated gene2)が、マウスパーキンソン病モデルおよび患者脳において活性化アストロサイトに強発現することを見出しました(Takeichi et al., 2011)。次に、NDRG2のノックアウトマウスを作製し、脳損傷モデルを用いて個体レベルでの解析を行った結果、NDRG2はIL-6/STAT3経路を介してアストロサイトの早期活性化に促進的に働いていることを発見しました(Takarada-Iemata et al., 2014)。また、脳梗塞モデルにおいても、NDRG2はアストロサイトによるグリア瘢痕形成や血液脳関門破綻の抑制を介して、梗塞巣の拡大に拮抗的な役割を持つことを観察しました (Takarada-Iemata et al., 2018)。一方、炎症性脱髄疾患である多発性硬化症のモデルにおいてNDRG2はむしろグルタミン酸輸送体の発現制御を介し神経症状の増悪に働くことを見出しました(Le et al., 2018)。
これらのことは、様々な病態下において、アストロサイトがNDRG2を介して神経細胞の生存や組織修復に多様かつ重要な役割を果たしている可能性を示しています。現在、更にアストロサイトがNDRG2を介しどのように病態形成に関与するのか、その分子機構の解明を行っています。
3.自閉症関連遺伝子による脳内代謝制御とグリアネットワークの形成
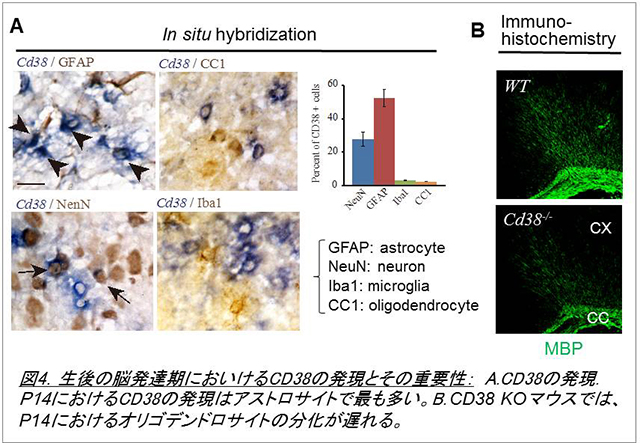
CD38は45kDaからなる膜蛋白質で、NAD+からcADPリボース、更にADPリボースを生成することが知られています。これまでの研究から、CD38ノックアウトマウスでは視床下部神経からのオキシトシン分泌量が低下し、母親の養育行動や社会的認識記憶に異常が認められることが明らかにされています(Jin et al., 2007)。このため、同マウスは自閉症モデルの一つと考えられています。一方、CD38は脳内でアストロサイトに最も強く発現していることから、私たちは特に生後発達期のグリア細胞及び神経細胞の発達にも影響を与えているのではないかという仮説を立てました。そして、野生型及びCD38ノックアウトマウスにおける生後のグリア発達、神経ネットワークの構築などについて検討を始めました。その結果、この時期においてもCD38は大脳皮質のアストロサイトで最も強く発現し、CD38ノックアウトマウスではアストロサイトの発達が遅延することを観察しました。更に興味深いことに、同マウスではオリゴデンドロサイトの分化も遅延することが明らかになりました(図4、Hattori et al., 2017)。そのメカニズムとして、基質であるNAD+が重要であることを発見しました。現在、アストロサイトに発現するCD38が脳内神経ネットワーク構築、更にマウスの行動にどのような影響を与えるか等について検討を行っています。
一方、CD38の欠損により引き起こされる脳内NAD+の増加はアルツハイマー病など病態モデルにおいては神経保護作用を持っていることが報告されています。今後、発達段階と病態下における脳内NAD+の役割の違いを明らかにできればと考えています。
4.末梢細胞による脳内炎症の誘導とその制御
脳内は血液脳関門を介して末梢組織から隔離されているため、通常は末梢の炎症が脳内には波及しない仕組みになっています。しかし、脳卒中などの病態ではこの関門が破壊され、白血球などの浸潤が認められます。私たちは、特にくも膜下出血や多発性硬化症の動物モデルを用いて、どのようにして末梢の炎症細胞が脳内に侵入し、脳内環境をどのように変化させるか等について解析を行っています。
5.ヒト脳の組織学的検討
金沢大学に御献体の提供をいただいた方のご家族の方へ
研究課題「ヒト脳の組織学的検討」について
近年,画像技術等の進歩により,ヒトの脳機能について新たな知見が多く得られてきております。たとえば,MRIの技術を応用することで,脳内の新たな機能局在やネットワークが明らかになってきました。一方,神経細胞やグリア細胞など、ヒトの脳を構成する細胞ごとの解析は,未だ限定的です。
本研究は,学生実習に使わせていただいた御献体の脳の一部を用いて,脳構成細胞および神経線維の同定を行うものです。実験動物等で得られた知見の検証,及びヒト神経病態との関連を検討する上で有用な染色法,及び抗体等が明らかになり,ヒトの脳研究の推進に貢献できると考えております。
本研究は,御献体いただいた方の解剖学の発展への助力という御遺志に沿う研究であり,患者様のご家族への新たな負担はありません。
なお,この研究は,金沢大学医学倫理審査委員会の審査を受け,研究域長の承認を得て行っているものです。
1.研究の対象
金沢大学に御献体を提供いただいた方の御家族で,この研究に参加したくない方がいらっしゃいましたら,そのことをお申し出ください。その場合,あなたの御家族の御献体の組織を研究に供することはありません。また,ご家族がこの研究の対象になっているかお知りになりたい方についても,お調べしお答えいたします。
2.研究の目的について
研究課題名::ヒト脳の組織学的検討
本研究では学生実習に使わせていただいた御献体の脳の一部を用いて,脳構成細胞および神経線維の同定を行います。これまで実験動物等で得られた知見の検証,及びヒトの疾患との関連を検討する上で有用な染色法,及び抗体等を明らかにすることを目的とします。
3.研究の方法について
この研究では学生実習に使わせていただいた御献体の脳の一部を用いて,脳構成細胞および神経線維の同定を試みます。具体的には,パラフィン包埋,切片作製の後に各種染色,免疫組織化学を行いて,神経細胞,グリア細胞,血管細胞等の脳構成細胞および神経線維の同定を行います。
情報を収集する前に,個人情報を完全に削除し個人を特定できないようにします。集計されたデータは学会や論文などに発表される事がありますが,個人情報が公表されることはありません。
4.研究期間
この研究の期間は,2025年2月21日(金沢大学医学倫理委員会の承認日)から2027年3月31日までです。
5.研究に用いる試料・情報の種類
試料:御献体の脳組織
6.外部への試料・情報の提供・公表
外部への試料・情報の提供はございません。上記の通り,集計されたデータは学会や論文などに発表される事がありますが,個人情報が公表されることはありません。
7.予想される利益と不利益について
この研究は御献体の脳組織を解析する研究であり,この研究に参加しても御家族に直接の利益はありません.予測される不利益として個人情報の流出の可能性が完全な0ではありませんが,データの取扱いについては,外部に漏れることが無いよう細心の注意を払います。また御献体の身体の組織の取り扱いに関しても,御家族が不快に思われるような扱いは決して致しません。
8.プライバシーの保護について
この研究では,研究に用いる組織に御献体に対応する番号をつけた一覧表を作り,データの調査には個人情報の含まれない対応番号のみを使います。また,上述の通り,この研究で得られた結果は学会や医学雑誌等に発表されることがありますが,御献体の個人情報などが公表されることは一切ありません。
9.研究参加に伴う費用の負担や通院について
この研究に参加することによる費用等の負担はありません。
10.研究組織
研究責任者:金沢大学 神経解剖学 教授 堀 修
研究分担者:金沢大学 神経解剖学 准教授 宝田 美佳
金沢大学 機能解剖学 教授 尾崎 紀之
金沢大学 総合技術部 技術専門職員 玉谷 貴志
11.研究への不参加の自由について
御献体が当該研究に用いられることについて,御家族・代理人の方にご了承いただけない場合には,研究対象としませんので,2026年3月31日までに下記の問い合わせ先までお申出ください。
12.研究計画書など資料の入手について
この研究の研究計画書などの資料が欲しい,またはごらんになりたい場合は,研究に関する窓口に問い合わせてくだされば,対応いたします。
13.研究に関する窓口
この研究の内容について,わからない言葉や,疑問,質問,自分がこの対象の対象になるかなど,更に詳細な情報をお知りになりたいときには,遠慮せずにいつでもお尋ねください。
研究機関の名称:金沢大学医薬保健研究域医学系
研究責任者:堀 修(金沢大学医薬保健研究域医学系教授)
問合せ窓口:堀 修(金沢大学医薬保健研究域医学系教授)
住所:金沢市宝町13-1
電話:Tel: 076-265-2162 Fax: 076-234-4222
研究代表者
金沢大学 堀 修
研究室で使用している疾患モデル動物(マウス)
| 脳梗塞 | 中大脳動脈閉塞モデル、両側総頚動脈閉塞モデル |
| パーキンソン病 | MPTP投与モデル、6-OHDA投与モデル |
| 頭部外傷 | Stab injuryモデル |
| 海馬神経細胞死 | カイニン酸投与モデル |
| 筋委縮性側索硬化症 | SOD G93Aトランスジェニックマウスモデル |
| 多発性硬化症 | MOG投与モデル |
| 緑内障 | 視神経損傷モデル |
| 自閉症 | CD38ノックアウトマウス |